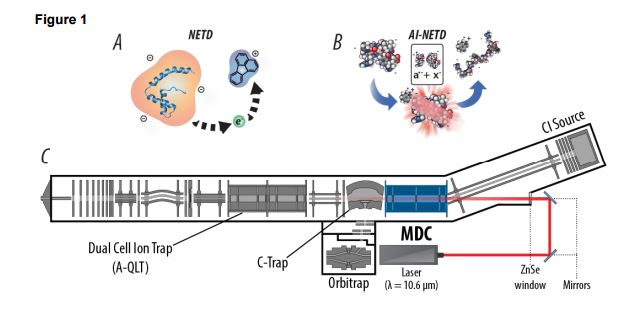
Whenever we talk about ideal dynamic exclusion settings I always start saying things like setting your numbers according to your peak widths. This week I realized that not everyone knows how to do this!
The way I do this is by opening a file in Xcalibur. Once I've done that I set some ranges with tight extraction windows so that I can look at a couple of good peptides. If its a human sample I look for two specific peptides. Their m/z(s) are:
722.32477
and
756.42590
These are two peptides from albumin. The first is relatively hydrophilic and the second is relatively hydrophobic. Since albumin is everywhere I'm going to always see them, whether its plasma or cell pellets or whatever.
To extract the peaks, click the thumbtack on your chromatogram. Next, right click anywhere on the chromatogram. Before you do anything else, toggle the tab over to the "Automatic processing" tab.
Set up the mass tolerance settings as above. I like 5ppm, though you can probably go lower on a recently calibrated Orbitrap, but 5ppm will do it. I don't use any smoothing or nuthin unless I'm trying to prepare a figure for people outside of the field to look at.
Next toggle back to ranges. You'll need to set it up as so:
So, add 2 ranges to your chromatogram tabs. Set them as base peaks and so that they only look at the MS1 scan filter. The box above shows how to search for the hydrophilic peptide. Repeat for the next row but enter the range(s) for the mass of the hydrophobic peptide. Hit "OK". You should now be looking at something like this:
You can't see much until you actually zoom in on the peak. You do this by clicking and dragging across the peak individually. On this 2 hour gradient the first one looks like this:
Now, we can just eyeball this and say "Yup! Thats about 30 seconds" or we can drag the mouse across the line. Normally in chromatography we would consider the peak width by measured across the peak at half height. This is NOT how we do this for dynamic exclusion. What you want to do is measure the peak from the intensity threshold cutoff in your method (or as close as you can get!)
(click to expand if you want to see it!
When this ion was first detected within this mass range on the instrument its intensity was 2e5. On a Q Exactive, my intensity threshold cutoff filter is typically in the 1-2e4 range. So the first time that this ion appeared in the chromatography it could be selected for fragmention. If it was then the dynamic exclusion would start counting at that point. In this case it was at 57.71 minutes. If I look at the far end of the peak it disappears around that range and it is just about exactly 0.5 minutes after it started. So this is a 30 second wide peak. Repeat for the second peptide.
It is important to keep in mind that some peaks will look different than others. In the case of these two peptides, I expect the second one to be wider than the first. From threshold to threshold on a 2 hour gradient it is 45s to 55s. Why is this? No idea. Not a chromatographer, but I can only set one value for my dynamic exclusion. If I'm a soloist I'm going to err for the higher value. If I'm a two-timer (
description of my terms for this here) I've got to be a little more intelligent and I might want to look at 4 or 5 more peaks to get a good idea of what my real peptide elutions peaks look like so I get the ideal number of MS/MS events.